

















Welcome to East of Eden Plants
Established by award-winning Botanist and Designer Peter Belton.
We are a professional team with an established working knowledge of:
Mediterranean, Hardy Exotic, Contemporary and Topiary Plants.
Plant propagation, Landscaping & Plantscape Installation.
We are a professional team with an established working knowledge of:
Mediterranean, Hardy Exotic, Contemporary and Topiary Plants.
Plant propagation, Landscaping & Plantscape Installation.
Sustainable Urban Greening & Biophilic Design including creation of
Natural Screening, Living walls, Green roofs, Rainwater Garden themes.
Commercial Interiors & Residential Garden Rooms,
Balcony, Terrace and Roof Garden Themes.
From Commercial and Residential projects to Private estates, Public and Community Spaces,
Our Inspirational themes and designs are created to be an expression of your
Lifestyle or Business Values.
Natural Screening, Living walls, Green roofs, Rainwater Garden themes.
Commercial Interiors & Residential Garden Rooms,
Balcony, Terrace and Roof Garden Themes.
From Commercial and Residential projects to Private estates, Public and Community Spaces,
Our Inspirational themes and designs are created to be an expression of your
Lifestyle or Business Values.









Expert Advice When You Need It
We are with you every step of the way. Our dedicated team of horticultural and landscaping experts are on hand to provide advice and guidance to help your achieve your vision.
Experienced plant experts are just a phone call away ready to discuss your planting requirements, the best plant species for your specific site, as well as essential planting and after care for vibrant foliage and healthy root growth. You will also receive comprehensive planting and care guides that have been curated by our team of experts to ensure your project runs as smoothly as possible for maximum results.
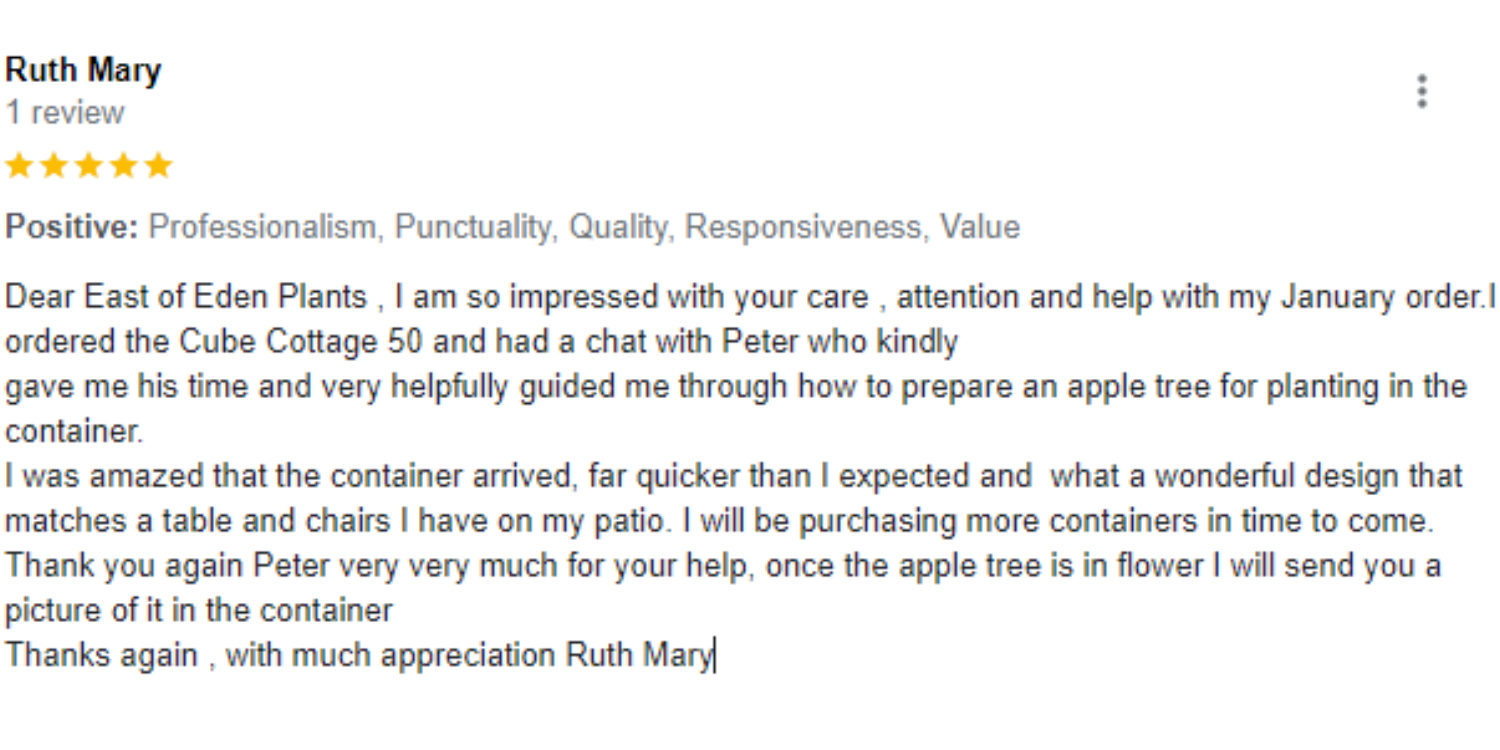
Don’t just take our word for it, here is what our clients had to say:
Don’t just take our word for it, here is what our clients had to say:




Menu Title
Contact Us
Send us your requirements - Click HERE

✕
Mailing List
Unsuccessful - please try again
Thank You
Thanks for subscribing to our mailing list.
Online - Start Chat?
Your name *